(第3回 院長通信)私の研究 その1
がんに対して、新しい治療標的の確立とそれに基づく創薬、がん免疫の活用と活性化、薬剤送達性の改善、陽子線、重粒子線といった粒子線療法、さらには早期診断や術前化学療法などさまざまなアプローチが世界中で試みられており、がんの治療成績は年々改善していますが、そのような中にあって、最も悪性である膵がんを筆頭に、難治がんの治療は依然として大きな課題となっています。
私は1978年神戸大学医学部の卒業で、1994年東北大学加齢医学研究所に着任、2024年までの30年間東北大学で研究生活を送りました。着任したときの研究テーマは、血管新生の分子メカニズムを解明し、その成果をもとにオリジナルな治療法を開発することでした。そして新規分子を探索することで2つの分子を発見し、2004年と2006年にそれぞれVasohibin-1 (VASH1)、Vasohibin-2 (VASH2)と命名して報告しました。VASH1は14番目の染色体長腕にコードされ、365個のアミノ酸で構成されるのに対し、VASH2は1番目の染色体長腕にコードされ、355個のアミノ酸で構成されています。
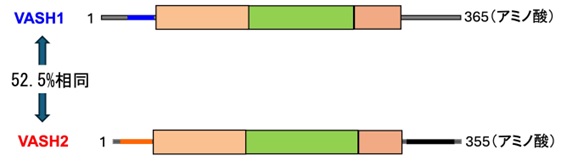
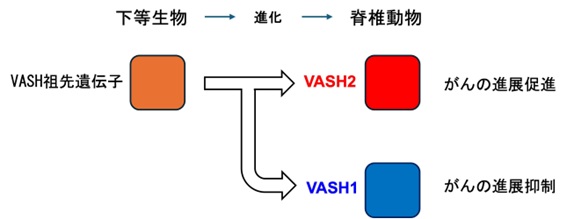
VASH1は、血管新生の刺激に応じて血管内皮細胞が産生し、血管新生を抑制するネガティブフィードバック因子として世界で初めて認定されました。これに対し、VASH2はVASH1とアミノ酸構造が50%以上一致していますが、VASH1とは反対に血管新生を促進することが明らかとなりました。さらに、VASHは種を超えて良く保存されていますが、血管の無い下等生物は単一のVASH祖先遺伝子を持ち、生物の進化に伴って血管を持つ脊椎動物からVASH1、VASH2の2つに別れたこと、塩基配列から見てVASH2の方がVASH祖先遺伝子により相同性が高いことが分かりました。つまり、もともと血管とは無関係に生物に付与されたVASHは、脊椎動物からは血管系にも使われるようになり、血管新生を促進するVASH2に対し、VASH1は逆に抑制する因子として分かれたと考えられるのです。
VASH1およびVASH2と各種病態との関連性について研究を進めました。そして、がん組織の病理学的解析からVASH1は腫瘍血管の内皮細胞に、VASH2はがん細胞に発現していることを明らにしました。さらに、さまざまな実験を通して、VASH1はがんの発育・転移を抑制するのに対して、VASH2はがんの発育・転移を促進することを実証することができたのです。